RNA編輯神器CasRx上線!shRNA要退休�����?
CRISPR/Cas系統(tǒng)賦予了細(xì)菌和古細(xì)菌適應(yīng)性免疫防御機(jī)制����,用于抵抗入侵的噬菌體及外源DNA。根據(jù)效應(yīng)蛋白的不同�����,CRISPR/Cas系統(tǒng)被分為兩類:第Ⅰ類�,依賴于多種效應(yīng)蛋白介導(dǎo)靶目標(biāo)干擾;第Ⅱ類����,以單一、多結(jié)構(gòu)域效應(yīng)蛋白為代表��。第II類CRISPR/Cas系統(tǒng)又分為II����、V和VI 3種類型。II型和V型效應(yīng)蛋白靶向于DNA分子����,例如II型CRISPR/Cas9系統(tǒng)����,憑借其低成本��、高效率和簡單易用等優(yōu)點(diǎn)�,成為DNA編輯的強(qiáng)大工具��,為基因操作和基因組研究帶來了革命性的改變。而VI CRISPR/Cas(CRISPR/Cas13)系統(tǒng)的獨(dú)特之處在于它們是目前發(fā)現(xiàn)的唯一能夠降解RNA的蛋白�����,CRISPR/Cas13系統(tǒng)的發(fā)現(xiàn)擴(kuò)展了CRISPR系統(tǒng)在病毒干擾方面的應(yīng)用�。新發(fā)現(xiàn)的Cas13蛋白第四種亞型(Cas13d)的家族成員CasRx在體內(nèi)外哺乳動物細(xì)胞的RNA裂解過程中顯示出更強(qiáng)的特異性和更高的敲除效率�,而且CasRx比之前發(fā)現(xiàn)的所有亞型都要小很多,更容易利用AAV載體實(shí)現(xiàn)體內(nèi)的運(yùn)輸��,因此在RNA編輯方面具有很大的應(yīng)用前景����。
1. CRISPR/Cas13系統(tǒng)
2016年��,科學(xué)家發(fā)現(xiàn)了可以靶向RNA進(jìn)行切割的CRISPR/Cas13系統(tǒng)����,該系統(tǒng)由單一的效應(yīng)蛋白Cas13和CRISPR RNA(crRNA)組裝形成一個(gè)由crRNA引導(dǎo)的RNA靶向效應(yīng)復(fù)合物�����。目前���,Cas13家族共鑒定出四種亞型�,包括Cas13a(又名C2c2)、Cas13b���、Cas13c和Cas13d�,四種Cas13亞型蛋白分子量均小于Cas9蛋白��。所有Cas13蛋白均具有兩種不同的催化活性:①由兩個(gè)較高等的真核和原核核苷酸結(jié)合(Higher eukaryotes and prokaryotes nuceotide-binding��,HEPN)結(jié)構(gòu)域提供的RNase活性,這是靶RNA降解所必需的�。Cas13 四個(gè)亞型同源性較低,且同源序列僅限于HEPN結(jié)構(gòu)域位點(diǎn)��,此外兩個(gè)HEPN結(jié)構(gòu)域中的單點(diǎn)突變會使CRISPR/Cas13系統(tǒng)對RNA的切割能力完全喪失��;②催化pre-crRNA加工和形成成熟crRNA的RNase活性。
圖1. VI CRISPR/Cas系統(tǒng)結(jié)構(gòu)示意圖
與CRISPR II類系統(tǒng)的其它核酸酶結(jié)構(gòu)一致�����,Cas13a的整體結(jié)構(gòu)是雙葉的����,N-末端結(jié)構(gòu)域(N-terminal domain����,NTD)和Helical-1結(jié)構(gòu)域構(gòu)成crRNA識別葉(REC lobe)����,HEPN1����、Helical-2、HEPN2結(jié)構(gòu)域形成核酸酶葉(NUC lobe)(圖2)����。CRISPR/Cas13b系統(tǒng)有兩種酶��,分別是VI-B1和VI-B2�����,它們之間的區(qū)別在于Cas13b轉(zhuǎn)座子上攜帶的附屬蛋白的基因型不同,VI-B1的附屬蛋白是Csx28�,而VI-B2的附屬蛋白是Csx27。絕大多數(shù)Cas13d型系統(tǒng)還含有包含 WYL結(jié)構(gòu)域的相關(guān)輔助蛋白���,WYL結(jié)構(gòu)域通常與原核防御系統(tǒng)有關(guān)(圖1)。
圖2. Cas13a蛋白結(jié)構(gòu)域示意圖
所有的Cas13蛋白酶都需要60- 66 nt的crRNA來確保靶向特異性���。四種亞型crRNA都有一個(gè)促進(jìn)與相應(yīng)Cas13蛋白酶結(jié)合的直接重復(fù)(Direct repeat��,DR)以及特異性靶向轉(zhuǎn)錄本的間隔序列。Cas13b的crRNA在3’端攜帶直接重復(fù)�����,而Cas13a�、c和d的crRNA在5’端攜帶直接重復(fù)(圖3)����。
圖3. 四種Cas13亞型對應(yīng)的crRNAs結(jié)構(gòu)示意圖
CRISPR/Cas13系統(tǒng)的防御機(jī)制主要包括以下幾個(gè)階段(以Cas13a為例):①pre-crRNA 的識別與結(jié)合�。新轉(zhuǎn)錄的pre-crRNA通過crRNA的5’端莖環(huán)結(jié)構(gòu)與Cas13a的REC葉識別并結(jié)合�,形成pre-crRNA與Cas13a復(fù)合物的中間過渡態(tài)��;②成熟crRNA的形成�����。NUC區(qū)的Helical-1和HEPN2結(jié)構(gòu)域之間保守殘基的構(gòu)象發(fā)生變化,從而形成一個(gè)酸堿催化中心���,催化酶切pre-crRNA形成成熟的crRNA�。此時(shí)的crRNA-Cas13a復(fù)合物處于無酶切活性狀態(tài)����;③crRNA-Cas13復(fù)合物酶切活性的激活。靶ssRNA進(jìn)入crRNA-Cas13a復(fù)合物內(nèi)與crRNA發(fā)生堿基互補(bǔ)配對�����,誘發(fā)Cas13a發(fā)生構(gòu)象變化�,從而激活crRNA-Cas13a復(fù)合物的酶切活性;④靶RNA的降解�。在crRNA的引導(dǎo)下��,Cas13a的HEPN結(jié)構(gòu)域催化靶ssRNA的酶切���。
有時(shí)細(xì)菌細(xì)胞中會出現(xiàn)非特異性酶切的情況�����,導(dǎo)致細(xì)胞中其他附屬ssRNA的降解��,引起一定的細(xì)胞毒性,但這種現(xiàn)象在哺乳動物細(xì)胞中并未出現(xiàn),其原因目前尚未可知�。(圖4)��。
圖4. CRISPR/Cas13系統(tǒng)作用機(jī)制示意圖
2. Cas13d的發(fā)現(xiàn)及優(yōu)勢
2018年����,加州大學(xué)伯克利分校Patrick D. Hsu實(shí)驗(yàn)室發(fā)現(xiàn)了靶向RNA的Cas13d家族�,Cas13d平均大小為930 aa,是目前發(fā)現(xiàn)的哺乳動物細(xì)胞中相對較小的Ⅱ類CRISPR效應(yīng)因子(比其家族成員Cas13a-c小20%����,比Cas9小33%)����,使它更容易包裝到容量有限的應(yīng)用載體如AAV載體中�����。此外���,Cas13d對RNA靶點(diǎn)的切割不依賴于PFS序列(protospacer flanking sequence,相當(dāng)于Cas9針對DNA的PAM序列)��,這極大的增加了Cas13d的應(yīng)用范圍���,使其成為進(jìn)一步開發(fā)靶向RNA工具的潛在平臺。與RNA干擾技術(shù)相比����,Cas13d介導(dǎo)的基因沉默具有更高的特異性(與數(shù)百個(gè)shRNA脫靶相比���,Cas13d沒有脫靶)和敲除效率(Cas13d達(dá)到96%����,shRNA達(dá)到65%�����, CRISPRi達(dá)到53%)�。而與Cas9介導(dǎo)的基因敲除技術(shù)相比����,Cas13d介導(dǎo)的基因沉默不會改變基因組DNA,因此這種基因沉默是可逆的��,對一些后天性疾病的治療更有優(yōu)勢(圖5��,圖6)�����。
圖5. 一種引導(dǎo)和靶向激活Cas13d核糖核酸酶活性的模型
圖6. Cas13d系統(tǒng)具有更高的特異性和敲除效率
與此同時(shí),Patrick D. Hsu實(shí)驗(yàn)室又鑒定出一種效果更佳的Cas13d酶:即來自腸道細(xì)菌黃化瘤胃球菌(Ruminococcus flavefaciens)XPD3002,被命名為CasRx����。隨后,科學(xué)家通過多項(xiàng)實(shí)驗(yàn)證明了CasRx能在體內(nèi)外可高效敲除RNA����。同時(shí)�,CasRx蛋白由于體積小,編輯效率高���,被認(rèn)為是在未來中極具應(yīng)用優(yōu)勢的Cas13蛋白��。
3. CasRx的敲除效果驗(yàn)證
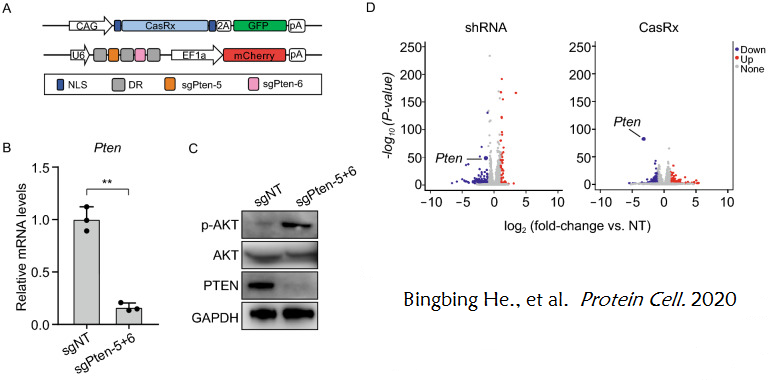
2020年的一項(xiàng)研究探索了使用CasRx系統(tǒng)來定向沉默基因的可行性。首先�����,研究者選擇Pten作為代謝調(diào)控基因�����,研究CasRx是否能以代謝基因?yàn)榘悬c(diǎn)進(jìn)行有效的基因敲除���,并制備了多個(gè)針對Pten mRNA編碼序列的sgRNAs,研究者將CasRx和每個(gè)Pten sgRNA通過質(zhì)粒轉(zhuǎn)導(dǎo)至小鼠神經(jīng)母細(xì)胞瘤N2a細(xì)胞中�����,并進(jìn)行sgRNA的體外篩選���,找到了CasRx敲低目的基因的最佳sgRNA組合:sgPten-5和sgPten-6。結(jié)果顯示�,在轉(zhuǎn)染CasRx���、sgPten-5和sgPten-6的N2a細(xì)胞中��,Pten的表達(dá)水平顯著降低�,AKT的磷酸化水平顯著升高�,此外與shRNA系統(tǒng)相比���,CasRx編輯系統(tǒng)表現(xiàn)出來更強(qiáng)的特異性和更低的脫靶率(圖7)。(延伸理解:Pten被認(rèn)為是一種代謝調(diào)節(jié)劑���,通過抑制PI3K/AKT途徑抑制胰島素信號轉(zhuǎn)導(dǎo)通路,Pten的缺失能促進(jìn)AKT的磷酸化)
圖7. CasRx體外介導(dǎo)Pten的敲除
研究人員通過尾靜脈注射敲低Pcsk9的AAV8病毒�,顯著降低了肝臟和血清中Pcsk9的表達(dá)量以及血清中的膽固醇水平����。重要的是,CasRx介導(dǎo)的Pcsk9敲低是可逆的�����,Pcsk9表達(dá)水平可以反復(fù)下調(diào)�。CRISPR/CasRx系統(tǒng)為可逆調(diào)節(jié)代謝基因,尤其是一些后天性疾?�。ㄈ缫虿涣忌盍?xí)慣導(dǎo)致的高血脂等后天代謝性疾病等)提供了一種有效的策略(圖8)�。
圖8. CasRx介導(dǎo)的血清膽固醇降低和PCSK9可逆調(diào)節(jié)
4. 維真生物推出RNA編輯新產(chǎn)品——CasRx
|
▽ 部分CasRx腺相關(guān)病毒(AAV)載體 ▽
|
|
pAV-CMV-nls-CasRx
|
pAV-CMV-nls-CasRx-P2A-GFP
|
|
pAV-U6-gRNA-CMV-nls-CasRx
|
pAV-U6-gRNA-CMV-nls-CasRx-P2A-GFP
|
▽ 實(shí)驗(yàn)檢測結(jié)果 ▽
維真科研團(tuán)隊(duì)針對mcherry基因設(shè)計(jì)了多條gRNAs,并選擇了其中兩條敲除效果好的gRNAs(gRNA1和gRNA2)��,進(jìn)行了單條gRNA、2in1 gRNAs敲除效果的驗(yàn)證試驗(yàn)����,結(jié)果如下:
從上述熒光圖可看出�,2in1 gRNAs敲除組��,mCherry表達(dá)水平大大降低�,敲除效果明顯好于單條gRNA的敲除效果�����。
(mcherry表達(dá)量:gRNA1組 0.38�����;gRNA2組 0.47����;gRNA1+2組 0.18)
此外���,從上圖mCherry mRNA表達(dá)量的檢測實(shí)驗(yàn)結(jié)果可以看出�����,Cas13d(CasRx)-gRNA系統(tǒng)可以特異高效地降低靶向基因的表達(dá)(mCherry)�����,并且2in1gRNAs 能實(shí)現(xiàn)mRNA水平敲低82%。
維真生物CasRx新品上線���,基因編輯效率更高,效果更好���,歡迎各位老師同學(xué)交流訂購����!
5. 文獻(xiàn)參考
[1]. Mahas, A., R. Aman and M. Mahfouz, CRISPR-Cas13d mediates robust RNA virus interference in plants. Genome Biol, 2019. 20(1): p. 263.
[2]. Huynh, N., et al., A versatile toolkit for CRISPR-Cas13-based RNA manipulation in Drosophila. Genome Biol, 2020. 21(1): p. 279.
[3]. He, B., et al., Modulation of metabolic functions through Cas13d-mediated gene knockdown in liver. Protein Cell, 2020. 11(7): p. 518-524.
[4]. Konermann, S., et al., Transcriptome Engineering with RNA-Targeting Type VI-D CRISPR Effectors. Cell, 2018. 173(3): p. 665-676.e14.
[5]. O'Connell, M.R., Molecular Mechanisms of RNA Targeting by Cas13-containing Type VI CRISPR-Cas Systems. J Mol Biol, 2019. 431(1): p. 66-87.
[6]. Liu, L., et al., The Molecular Architecture for RNA-Guided RNA Cleavage by Cas13a. Cell, 2017. 170(4): p. 714-726.e10.